產(chǎn)單核細胞李氏桿菌噬菌體分離、超離、宿主譜鑒定及生長曲線繪制(二)
2結(jié)果
2.1產(chǎn)單核細胞李氏桿菌噬菌體的分離
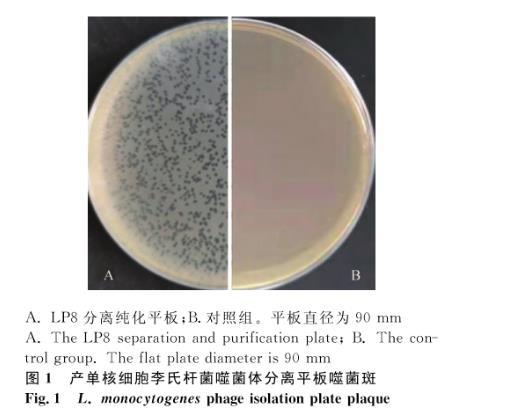
利用本實驗室保存的產(chǎn)單核細胞李氏桿菌作為宿主菌,對采集的所有污水樣品利用雙瓊脂平板法進行分離,分離出若干株噬菌體,選取一株性狀優(yōu)良、傳代穩(wěn)定的噬菌體進行后續(xù)試驗,并命名為LP8,宿主菌編號為ATCC19111(1/2a),平板上可以觀察到明顯的噬菌斑,噬菌斑形狀規(guī)整,邊緣整齊,顏色透亮,直徑2 mm左右,純化傳代穩(wěn)定,無暈環(huán)(圖1)。
2.2噬菌體超離
利用氯化銫密度梯度離心法對LP8噬菌體濃縮液進行超離,在密度為1.50~1.70 g·mL的氯化銫溶液梯度之間具有噬菌體層,吸取噬菌體超離液,通過效價核定,效價為4.4×10PFU·mL。
2.3噬菌體電鏡觀察
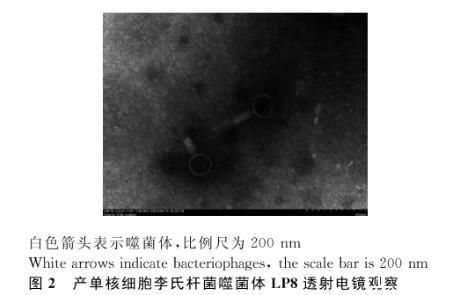
利用透射電鏡對LP8樣品進行觀察,發(fā)現(xiàn)LP8為肌尾科噬菌體,具有標(biāo)準(zhǔn)的20面體的頭部,還有一個由中空的針狀結(jié)構(gòu)和鞘組成的尾部,以及尾針組成的基部,噬菌體的頭部直徑約為80 nm;尾部長度約為120 nm(圖2)。
2.4 LP8噬菌體效價的測定
對LP8噬菌體的裂解液、濃縮液、超離液進行效價測定,結(jié)果分別為1.76×10、2.67×10、4.4×10PFU·mL。
2.5 LP8噬菌體宿主譜鑒定
利用實驗室所保存的產(chǎn)單核細胞李氏桿菌、威爾斯李氏桿菌和英諾克李氏桿菌對LP8噬菌體進行宿主譜鑒定,結(jié)果顯示LP8噬菌體對18株LM都具有裂解性,對5株威爾斯李氏桿菌(C10、9-2、4-40、C59、4-43)具有裂解性,對22株英諾克李氏桿菌均無裂解性(表1)。
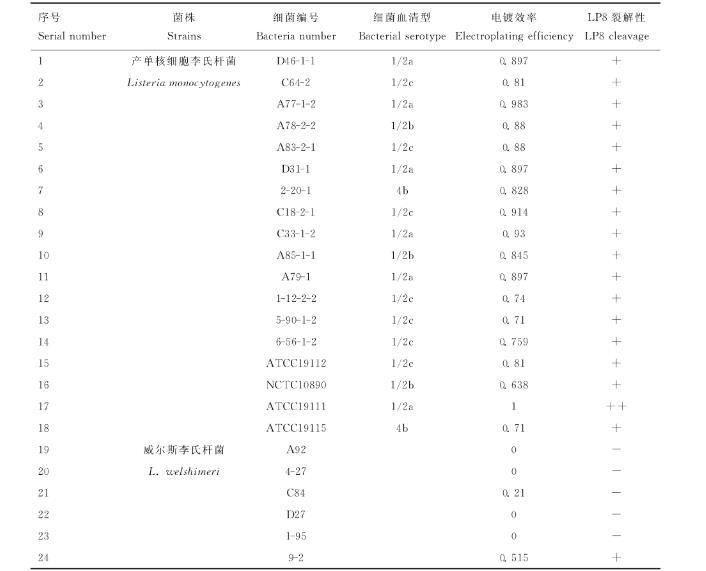
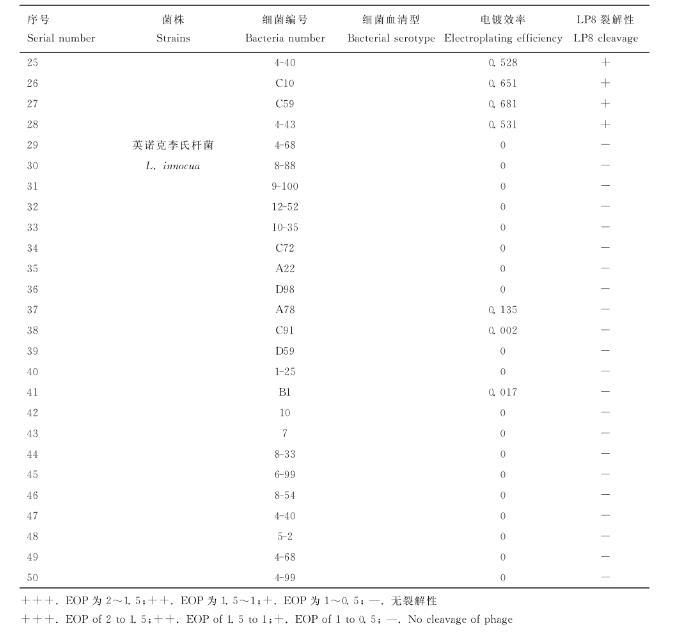
表1 LP8噬菌體宿主譜
2.6 LP8噬菌體生長特性
對LP8設(shè)置不同的感染比例,測定出LP8的MOI為1(圖3A);設(shè)置25、35、45、55、65、75℃溫度梯度,確定LP8的最適溫度為45℃(圖3B);設(shè)置pH為2、4、6、7、8、10、12梯度,確定LP8的最適pH為7(圖3C);采用的6種有機溶劑中,氯仿和異戊醇對LP8影響最大,可造成LP8全部失活,丙三醇和SM空白對照組沒有影響,甲醇和乙醇會導(dǎo)致噬菌體部分失活(圖3D);繪制一步生長曲線,確定LP8潛伏期為23 min,對數(shù)期為23~80 min,80 min以后達到平臺期,噬菌體效價最大可達到3.4×10PFU·mL(圖3E)。
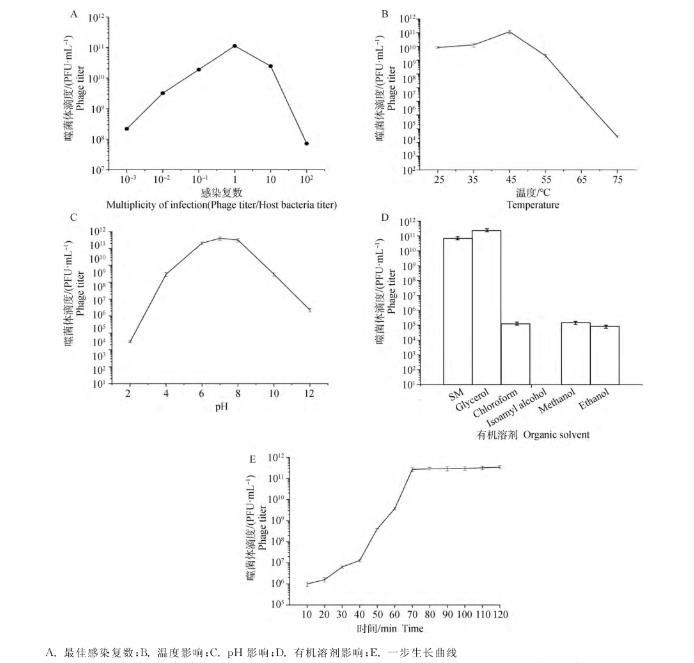
A.最佳感染復(fù)數(shù);B.溫度影響;C.pH影響;D.有機溶劑影響;E.一步生長曲線
2.7 LP8基因組酶切鑒定
電泳條帶顯示,噬菌體LP8基因組在1、2號DNA酶泳道未出現(xiàn)條帶,在3、4號RNA泳道和5、6號綠豆核酸酶泳道都出現(xiàn)了條帶,根據(jù)條帶結(jié)果得出噬菌體基因組為雙鏈DNA(圖4)。
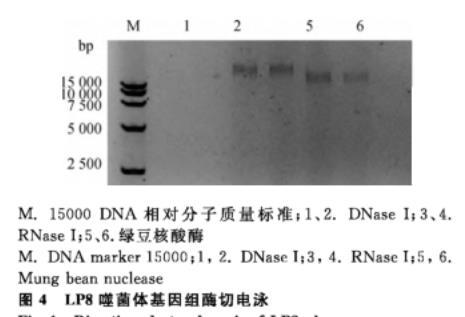
M.15000 DNA相對分子質(zhì)量標(biāo)準(zhǔn);1、2.DNase I;3、4.RNase I;5、6.綠豆核酸酶
2.8 LP8基因組測序
經(jīng)測序,噬菌體LP8的基因組為87 038 bp,共有120個編碼基因,編碼基因的累加長度為76 326 bp、編碼基因的平均長度為636 bp、編碼區(qū)域長度占基因組的比例為87.69%。通過SnapGene軟件對全基因組進行繪圖,分析其基因的大小和分布情況。通過COG、KEGG、GO對LP8的基因進行注釋,發(fā)現(xiàn)有6個編碼基因參與原噬菌體合成;有20個編碼基因參與噬菌體有關(guān)的生物過程;11個編碼基因參與分子功能,如結(jié)合或催化;8個編碼基因參與噬菌體的合成,具體生物學(xué)通路最多的為嘌呤代謝和嘧啶代謝(圖5)。
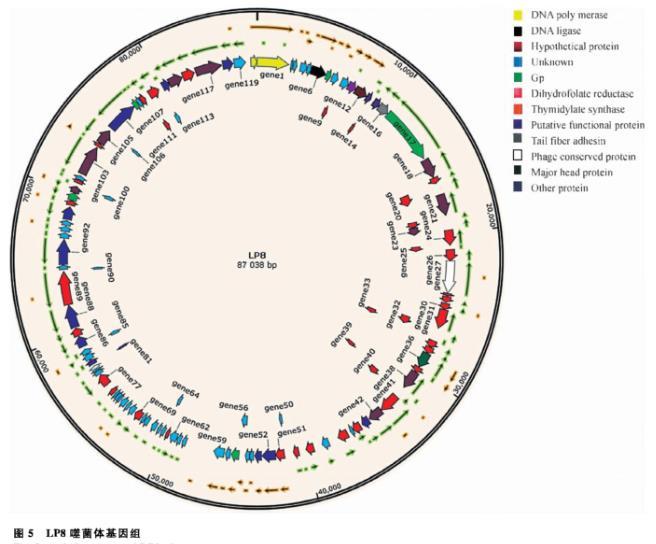
圖5 LP8噬菌體基因組
相關(guān)新聞推薦
2、高效乳酸菌和纖維素分解菌混合培養(yǎng)是否會相互抑制生長?
3、腸道微生物代謝物丁酸鹽可維持腸道內(nèi)穩(wěn)態(tài)和調(diào)節(jié)腸道炎癥