微生物合成食品功能因子方法、研究策略和進展(一)
隨著社會發(fā)展和人們對安全、健康、營養(yǎng)飲食的追求,功能性食品的研發(fā)成為食品領(lǐng)域的研究熱點[1].其中,食品功能因子是功能性食品制造的基本原料,也是賦予食品特定生理功能的關(guān)鍵成分.食品功能因子種類豐富[2],根據(jù)其化學(xué)結(jié)構(gòu)主要分為11類:活性多糖,功能性低聚糖和功能性單雙糖,功能性脂類,糖醇類,多糖類,氨基酸、肽與蛋白質(zhì),維生素和維生素類似物,礦物元素,植物活性成分,益生菌和低能量食品成分.這些功能因子在調(diào)節(jié)人體機能,改善睡眠和促進生長發(fā)育等方面發(fā)揮重要作用.
合成生物學(xué)作為一種更安全、更健康和綠色可持續(xù)的食品獲取方式,為功能性食品組分和重要功能營養(yǎng)因子的生產(chǎn)提供了技術(shù)支撐,成為推動食品行業(yè)發(fā)展的重要手段.以大腸桿菌(Escherichia coli,E.coli)、枯草芽孢桿菌(Bacillus subtilis,B.subtilis)、谷氨酸棒桿菌(Corynebacterium glutamicum,C.glutami cum)和釀酒酵母(Saccharomyces cerevisiae,S.cerevi siae)等為代表的典型模式微生物具有生長速度快、營養(yǎng)要求簡單、遺傳操作簡便和工業(yè)發(fā)酵培養(yǎng)模式相對成熟等優(yōu)勢[3,4],已廣泛用于合成各種生物能源和生物醫(yī)藥等[5~7],同時在食品領(lǐng)域也被開發(fā)用于高效合成各種營養(yǎng)組分,如N-乙酰氨基葡萄糖[8,9]、母乳中的關(guān)鍵成分2′-巖藻糖基乳糖[10]、血紅蛋白[11]以及乳蛋白[12]等.其中,細胞的生長調(diào)控是實現(xiàn)高效生物制造的關(guān)鍵控制點,因此以模式微生物底盤細胞作為食品功能因子的合成載體,充分利用合成生物學(xué)原理和技術(shù)對細胞的生長進行有效精細調(diào)控,有望實現(xiàn)食品功能因子的高效合成.本文主要從轉(zhuǎn)運蛋白工程改造提高菌株生長速率、重編程細胞能量代謝、細胞生長與產(chǎn)物合成偶聯(lián)以及細胞生長與產(chǎn)物合成解偶聯(lián)四個方面系統(tǒng)總結(jié)了目前基于模式微生物生長調(diào)控合成食品功能因子的研究策略和進展(圖1),并指明微生物合成食品功能因子面臨的挑戰(zhàn)及未來發(fā)展方向,以期為功能因子的高效生物制造提供新思路.
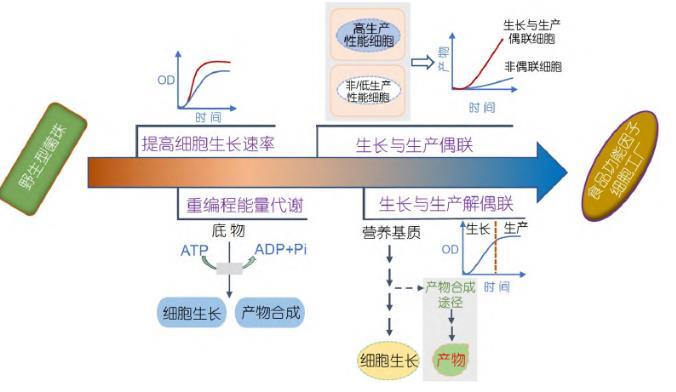
1優(yōu)化底盤細胞生長速率
發(fā)酵過程中微生物有效吸收和代謝關(guān)鍵營養(yǎng)基質(zhì)的能力直接影響細胞的生長.因此,提高碳源利用率或擴大可利用碳源的范圍能夠優(yōu)化底盤細胞的生長速率,增加細胞密度,從而實現(xiàn)食品功能因子的高效合成[13].
1.1碳源轉(zhuǎn)運蛋白工程
微生物可以利用多種碳源,它們對碳源的選擇主要取決于胞內(nèi)相關(guān)碳源轉(zhuǎn)運蛋白的表達,因此高效表達碳源轉(zhuǎn)運蛋白是提高碳源利用率的一種直觀快速方法.研究較為廣泛的是S.cerevisiae[14~18]和E.coli[19~23]中的各種內(nèi)源和外源糖類轉(zhuǎn)運蛋白.S.cere visiae對己糖特別是葡萄糖的轉(zhuǎn)運機制已被很好地表征,它通過內(nèi)源的Hxt家族己糖轉(zhuǎn)運蛋白介導(dǎo)的轉(zhuǎn)運系統(tǒng)來利用己糖[18,24].目前,已鑒定出約20種編碼己糖轉(zhuǎn)運蛋白的Hxt基因,研究表明,通過調(diào)節(jié)一種或幾種轉(zhuǎn)運蛋白的表達可提高細胞對葡萄糖的攝取能力,以S.cerevisiae CEN.PK m850作為出發(fā)菌株,經(jīng)過改造使其具備乳酸合成能力,乳酸的生產(chǎn)強度為0.77 g/(L h),后通過強化HTX1基因的表達,提高菌株對葡萄糖的消耗速率,使得乳酸生產(chǎn)強度達到(0.85±0.02)g/(L h)[25].考慮到發(fā)酵成本和環(huán)境保護等因素,越來越多的糖組分成為微生物可利用的替代碳源[26].在無特異性轉(zhuǎn)運蛋白存在時,添加非天然碳源會明顯減弱底盤細胞的生長速率.而Wang等人[27]通過引入低親和力的阿拉伯糖轉(zhuǎn)運蛋白araE基因,使得S.cere visiae在以蔗糖為碳源的培養(yǎng)基中高效合成白藜蘆醇,產(chǎn)量達2.3 mg/L.因此,研究人員可通過引入外源轉(zhuǎn)運蛋白或改造內(nèi)源Hxt轉(zhuǎn)運蛋白,使得S.cerevisiae在以蔗糖、果糖或纖維二糖等非天然碳源為營養(yǎng)基質(zhì)的條件下進行有效生長,從而賦予其高產(chǎn)果聚糖[28]、檸檬酸[29]等食品功能因子的能力.Bruschi等人[30]在不能以蔗糖為碳源的E.coli基因組上整合表達蔗糖利用基因表達盒,發(fā)現(xiàn)菌株以蔗糖為碳源和以葡萄糖為碳源時的生長速率一致,這為以蔗糖或含蔗糖的廉價原料為碳源有效合成產(chǎn)品提供了可能.一方面,通過調(diào)節(jié)轉(zhuǎn)運蛋白表達強度或繼續(xù)挖掘其他碳源轉(zhuǎn)運蛋白有利于提高碳源攝取率并擴大可利用碳源種類,從而改善底盤細胞的生長性能和生產(chǎn)潛力.另一方面,轉(zhuǎn)運蛋白可能受到轉(zhuǎn)錄因子及其效應(yīng)物的調(diào)控[31,32].因此,深入了解其調(diào)控機制可為合成食品功能因子底盤細胞的改造提供理論依據(jù).
1.2適應(yīng)性實驗室進化
適應(yīng)性實驗室進化(adaptive laboratory evolu tion,ALE)是一種在特定生長環(huán)境或選擇壓力下,通過長期培養(yǎng)篩選獲得具有某種更優(yōu)特性菌株的技術(shù).它無需深入了解細胞錯綜復(fù)雜的代謝網(wǎng)絡(luò),只需根據(jù)目標表型設(shè)計選擇壓力來篩選獲得較優(yōu)突變株,具有適用范圍廣、實用性強等優(yōu)點[33~35].適應(yīng)性實驗室進化已被廣泛用于優(yōu)化微生物底盤細胞的生長表型,以改善菌株的生產(chǎn)性能[36~39].B.subtilis作為GRAS(generally recognized as safe)菌株,已廣泛用于高效合成多種功能性低聚糖[40,41]、功能性單糖[42]及蛋白質(zhì)[43]等食品功能因子.Liu等人[44]將野生型B.subtilis 168在M9基本培養(yǎng)基中傳代超過1000代,進化菌株A40的比生長速率提升43.5%,乙偶姻和卵清蛋白生物合成分別提高54.5%和20%.多層次組學(xué)技術(shù)可用于闡明進化株的進化機制及其基因型和表型之間的關(guān)聯(lián),后者可用于指導(dǎo)構(gòu)建或優(yōu)化高效合成目標產(chǎn)物的細胞工廠.LaCroix等人[45]對E.coli MG1655進行為期81天的進化后獲得一株生長速率提高1.6倍的菌株.全基因組測序發(fā)現(xiàn),主要是pyrE/rph基因間隔區(qū)、全局轉(zhuǎn)錄基因rpoB和DNA結(jié)構(gòu)基因hns/tdk間隔區(qū)發(fā)生突變,pyrE/rph突變與菌株的嘧啶生物合成相關(guān).經(jīng)轉(zhuǎn)錄組學(xué)分析,rpoB和hns/tdk突變導(dǎo)致與蛋白合成、葡萄糖轉(zhuǎn)運和糖酵解途徑相關(guān)基因的表達水平上調(diào),增加中心碳代謝途徑的通量,從而提高底盤細胞的生長速率.研究表明,C.gluta micum在以葡萄糖為唯一碳源的基本培養(yǎng)基中經(jīng)過1500代培養(yǎng)后,其生長速率提高了42%.經(jīng)基因組測序和反向代謝工程發(fā)現(xiàn),轉(zhuǎn)錄調(diào)控因子GntR1和RamA的突變導(dǎo)致菌株生長性能的改善[46].因此,適應(yīng)性實驗室進化結(jié)合組學(xué)分析可鑒定出提高底盤細胞生長適應(yīng)性的關(guān)鍵突變點,有助于實現(xiàn)食品功能因子的高效生物合成.
適應(yīng)性實驗室進化也可被用于優(yōu)化微生物對碳源的攝取率,或者擴大微生物對替代碳源,如木糖、乙酸鹽和檸檬酸鹽等的利用范圍以提高細胞生長速率[34,47].木糖作為木質(zhì)纖維素中含量第二豐富的糖類,如何高效轉(zhuǎn)化為功能營養(yǎng)品等高附加值產(chǎn)品是研究熱點.Promdonkoy等人[48]在以木糖作為唯一碳源的培養(yǎng)基中對S.cerevisiae進行適應(yīng)性實驗室進化,提高了其對木糖的攝取能力,并鑒定出一株生長速率提高58%且異丁醇產(chǎn)量提升110%的菌株.Seong等人[49]以E.coli DSM01作為出發(fā)菌株,在以乙酸鈉為唯一碳源的M9培養(yǎng)基中傳代培養(yǎng)4 0天后獲得一株生長速率為0.14 h?1的進化株,而DSM01在該培養(yǎng)條件下幾乎無法生長.隨后驗證該進化菌株合成萜類化合物中間體甲羥戊酸(mevalonate,MVA)的能力,MVA的生產(chǎn)強度達到1.32 g/(L h),相比出發(fā)菌株提高14.3倍,這加速了乙酸鹽作為碳源生產(chǎn)功能營養(yǎng)品、生物燃料和重組蛋白等的研究進展.適應(yīng)性實驗室進化也可用于實現(xiàn)微生物對混合碳源的吸收利用.例如,Reider Apel等人[16]對引入木糖利用途徑的S.cerevisiae BY4742菌株以木糖為碳源進行培養(yǎng)進化,進化后菌株經(jīng)全基因組測序,發(fā)現(xiàn)己糖轉(zhuǎn)運蛋白HXT7(F79S)發(fā)生突變,該突變一方面提高了S.cerevisiae在以木糖為唯一碳源時對木糖的攝取率,從而展現(xiàn)出顯著的生長;另一方面,在存在雙碳源——葡萄糖和木糖時顯示出可同時利用兩種碳源的優(yōu)良特性,從而減少培養(yǎng)時間并提高生產(chǎn)效率.由此可見,適應(yīng)性實驗室進化可以有效提高微生物對不同底物的利用率和對環(huán)境的適應(yīng)性,進化得到的高生長速率底盤細胞在改善產(chǎn)物合成方面具有很好的潛力(表1).
適應(yīng)性實驗室進化與合成生物學(xué)和系統(tǒng)生物學(xué)工具相結(jié)合是一種優(yōu)化菌株表型和性能的有效策略,刪除有害突變、保留有益突變可重構(gòu)性能優(yōu)異的微生物底盤細胞.適應(yīng)性實驗室進化通過非直觀的方式提高微生物的適應(yīng)性,但是由于突變本身很難達到精準預(yù)測,同時較長的進化周期可能導(dǎo)致錯失獲得最優(yōu)表型的突變株.而基因組規(guī)模代謝模型[62~64]可揭示表型與基因型的相關(guān)性并用于預(yù)測生理變化,兩者相互結(jié)合補充可以更好地用于優(yōu)化菌株表型或開發(fā)新的微生物平臺.
相關(guān)新聞推薦
2、拮抗菌生長曲線測定、鑒定及對煙草赤星病室內(nèi)防治效果(一)
3、微生物發(fā)酵飼料原料來源、常用菌種及生長條件、應(yīng)用