北京及非北京基因型結核分枝桿菌的生長曲線與基因表達差異
1995年,在中國北京及周邊地區首次發現并報道MTB北京基因型菌株,將其命名北京型菌株。全球MTB臨床分離株的13%是北京基因型菌株,并導致了全世界1/3的TB。與非北京基因型菌株相比,北京基因型菌株具有一定的選擇優勢,傳播速度更快,毒力更強,往往更加致命。究竟什么原因賦予北京基因型菌株的這些特點還不完全清楚,被認為是相關特定分子的內在表達變化所決定。
研究結果顯示,北京和非北京基因型菌株其生長曲線總體趨勢都是標準條件下的OD值高于低氧及營養饑餓條件,正常標準培養條件下,由于氧氣、營養充足、充分,適合MTB生長繁殖,OD值高;低氧條件下OD值低,MTB生長輕度受限,這種不利條件也是促進MTB持留生存的因素之一;營養饑餓條件下OD值最低,說明MTB生長速度最慢,繁殖速度降低以利于存活,也許這與MTB入侵人體被巨噬細胞吞噬后不被清除變為休眠菌機制相一致。與非北京基因型菌株相比,北京基因型菌株在低氧、營養饑餓的條件下OD值均高,即在不利條件下,北京基因型菌株生長繁殖速度快于非北京基因型菌株,說明其對低氧、營養饑餓不利條件耐受性較好,在壓力條件下生存能力強,這也許是北京基因型菌株易流行的原因之一。是MTB什么內在的因素造成了這種不同,作者初步探索了MTB菌株的毒素-抗毒素mazEF系統與其生長繁殖的相關性。
毒素-抗毒素(TAS)由毒素蛋白和同源的抗毒素組成,mazEFTAS能夠促進程序性細胞死亡,抑制細菌生長,誘導持留菌的形成,造成潛伏感染及提高細菌對環境壓力的耐受性。TAS的作用機制取決于病原菌在不同生活時期的生物學特性,當受到不良環境或刺激后,打破了毒素抗毒素之間的平衡,抵抗力強的毒素被激活,作用于胞內靶分子,引起抑菌或殺菌效應。
Sat等對大腸桿菌進行mazEF基因敲除,通過比較其普通菌株和敲除株對不利環境如藥物的反應,發現藥物可以干擾或消耗不穩定的抗毒素,而使穩定的毒素MazF堆積,導致細菌的程序性死亡,因此推測mazEF可能是其程序性死亡進程中的關鍵因素。本研究通過敲除和過表達北京和非北京基因型菌株mazEF3、mazEF6和mazEF9基因后,觀察各菌株在不同環境下的生長曲線,結果顯示無論是北京還是非北京基因型菌株,其缺失突變株和過表達菌株的OD值均低于親本株,提示mazEF3,6,9與MTB生長繁殖相關。缺失后,mazEF3,6,9不能發揮其應有的調節細菌生長繁殖作用,尤其是在不利的環境條件下,導致MTB菌數下降。過表達mazEF3、mazEF6和mazEF9基因,MTB菌數也降低,推測雖然抗毒素可以中和毒素,但是抗毒素不穩定,易降解,導致毒素增多、引起菌體死亡,或者是由于菌體內其他調節因素也參與了對其過表達調節作用的結果。究竟是何種原因導致的這種現象,需要進一步探討。該結果也提示作者應進一步研究:在上述培養條件下,分別檢測mazE3、mazE6、mazE9、mazF3、mazF6和mazF9對北京和非北京基因型菌株生長繁殖的影響。在mazEF系統mRNA表達水平上,北京基因型與非北京基因型菌株相比,mazEF3高表達;mazF3、mazF6、mazF9均高表達;mazE9低表達,差異均具有統計學意義(P<0.05)。
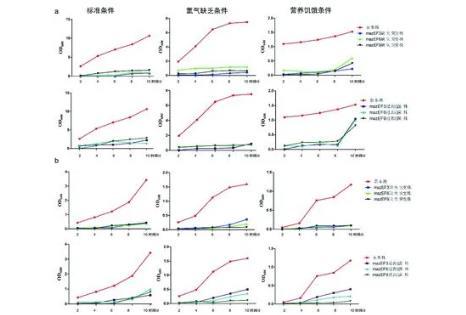
注:a. 北京基因型菌株及其缺失和過表達菌株在不同培養條件下的生長曲線;b. 非北京基因型菌株及其缺失和過表達菌株在不同培養條件下的生長曲線。圖2 結核分枝桿菌及其mazEF3,6,9基因缺失和過表達菌株的生長曲線
有研究表明,分枝桿菌中Rv1103c-1102c(mazEF3)、Rv1991a-1991c(mazEF6)和Rv2801a-2801c(mazEF9)屬于經典的TAS,其中mazEF6、mazEF9在營養匱乏環境中啟動子增強最顯著。本研究結果顯示北京基因型比非北京基因型菌株的mazEF3基因轉錄水平高,兩者同為臨床致病菌,或許mazEF3高表達賦予了北京基因型菌株抗壓力強的特點,在入侵宿主時可以抵抗機體的不利環境而生存,這也許是造成其易流行、持留生存及出現耐藥菌性的原因之一。當MTB處于不利環境時或入侵宿主遭遇免疫系統攻擊時,接受細胞外死亡因子的刺激,細菌的TAS被激活,其中毒素性質穩定,而抗毒素性質不穩定易被蛋白水解酶水解,造成毒素累積,而北京基因型菌株毒素基因均高表達,可產生更多的毒素,可以選擇性的使一些細菌死亡,從而為剩余的細菌保留能量和營養物質,此為利他性死亡,以保證群體穩態和適應不利環境。
作為新疆主要流行株北京基因型菌株mazE9低表達,推測為是內在特定分子表達調節的結果,尤其是mazE9低表達和mazF9高表達的協同作用,能使更多的毒素累積進行利他性程序性死亡,從而提高自身對不良環境的耐受性,也許這也是成為流行株的原因之一。
研究證實大量誘導表達MazF可使細胞處于停滯狀態,隨后大劑量誘導表達MazE可中和MazF的毒性,使細胞恢復活性。這一停滯狀體與結核分枝桿菌的休眠狀態很相似。當游離的MazF在細胞內時間或累計量超過某一界點時,就會導致細胞的死亡。我們推測毒素蛋白累積量不同可以使細胞表現出不同的狀態,當少量累計時提高細胞對不利生存環境的耐受,當環境進一步惡化,毒素持續累積,促使細胞進入休眠狀態,當毒素作用時間或者累積量達到某一值時細胞死亡。實驗中使用Western Blot技術檢測蛋白質水平的表達時,結果顯示,北京基因型菌株相對于非北京基因型菌株,MazF9蛋白高表達,與mazF9基因mRNA表達趨勢一致。在轉錄和翻譯水平上,mazF9的高表達也許是北京基因型菌株成為流行株、廣泛傳播的原因之一。
綜上所述,作者初步探索了北京及非北京基因型結核分枝桿菌的生長曲線、mazEF3,6,9系統表達有無差異以及mazEF系統是否與北京基因型菌株流行相關。研究結果表明,北京與非北京基因型結核分枝桿菌的生長曲線、mazEF3,6,9系統表達存在差異,mazEF系統與北京基因型菌株流行相關。MTB的染色體上存在著大量的TAS系統,mazEF3,6,9系統只是其中之一,mazEF3,6,9系統在MTB中究竟如何發揮作用以及與其他TAS系統、調節因素的互作用需要進一步研究。
相關新聞推薦
1、產脂肪酶地衣芽孢桿菌LD-1302篩選、培養基及產酶條件研究(一)